
陆生爬行动物和第一批哺乳动物
在陆地上,三叠纪的脊椎动物以迷宫两栖动物和爬行动物为代表,后者由后肢科,节肢动物,风生动物,科齿科动物和变形龙组成。在二叠纪末期,所有这些四足动物群的多样性都急剧下降。75%的早期两栖动物家庭和80%的早期爬行动物家庭消失在二叠纪-三叠纪边界或附近。早期的三叠纪形式仍然是古生代的,而新的形式在整个时期都出现了,到三叠纪晚期,四足动物区系显然是中生代的。在三叠纪中晚期首次出现祖先形态的现代群体包括蜥蜴,乌龟,鼻头动物(蜥蜴类动物)和鳄鱼。
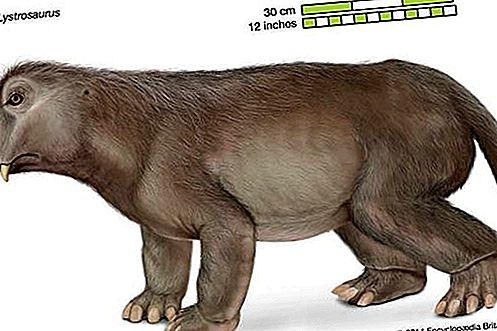
象哺乳动物一样的爬行动物或类兽类在二叠纪晚期遭受了灭绝的冲击。该组织在边界危机中幸存下来,但到三叠纪末期实际上已灭绝,这可能是由于来自更有效率的掠食者(如thecodonts)的竞争。该类动物中最著名的一种是Lystrosaurus,其化石已在印度,南部非洲和南极洲发现。因此提供了证明这三个大陆曾经联系在一起的证据。
最初很小的真正的哺乳动物出现在三叠纪晚期(例如,类似Morgan的Morganucodon)。尽管它们的化石遗骸是从三叠纪末的Rhaetian阶段的英国骨床上收集的,但是保存完好的化石并没有清楚地表明从三叠纪末期从爬行动物到哺乳动物的进化过渡。
第一批恐龙
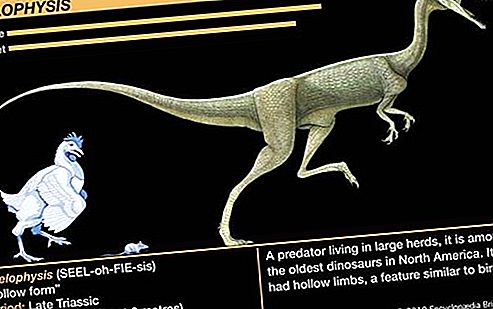
齿科最早在三叠纪早期遇到,在中三叠纪期间很常见,但在侏罗纪开始之前就消失了。在三叠纪中,这组考古学家(或“统治爬行动物”)中典型的是属于伪类动物的小型两足动物。象Lagosuchus这样的形式是快速运行的捕食者,它们在身体的正下方竖立着四肢,这使它们更加灵活机动。在三叠纪晚期至侏罗纪早期,这个群体大概产生了属于saurischian和ornithischian阶的原始恐龙。早期的恐龙是双足的,活动迅捷,与后来的中生代恐龙相比相对较小,但是有些恐龙,例如板龙(见图),长度达到8米(26英尺)。腔骨龙(见图)是一种三叠纪晚期食肉恐龙,长约2米(6至8英尺)。它的化石是在美国亚利桑那州东北石化森林国家公园的Chinle组中发现的。恐龙群后来在中生代变得更为重要,因此这个时代被非正式地称为“爬行动物时代”。
飞行的爬行动物
一些最早的蜥蜴可能是最早登上空中的脊椎动物。滑翔的蜥蜴,例如小型三叠纪的伊卡龙,被认为是从伸展的肋骨之间伸展的皮肤上形成了翼型,这将使短距离的滑行与当今的松鼠产生的滑行相似。同样,Longisquama具有可以用作原始翅膀的长鳞片,而三叠纪晚期Sharovipteryx是活跃的飞行者,可能是第一个真正的翼龙(飞行爬行动物)。所有这些形式在三叠纪末期都灭绝了,它们的飞行角色被后来的侏罗纪和白垩纪的翼龙所取代。
植物
陆生植物受二叠纪-三叠纪危机的影响,但受动物影响的程度却不及动物,因为晚古生代植物群的灭绝已经开始。三叠纪主要的底层植物是蕨类植物,而大多数中层植物是裸子植物(裸露种子的植物)—苏铁类(已灭绝)和苏铁类和银杏类。三叠纪森林的高层由针叶树组成。他们最著名的化石遗迹保存在上三叠纪Chinle组中。
虽然在三叠纪确实存在广泛的森林,但是在三叠纪早期和中三叠纪北部大陆的广泛干旱限制了它们的面积范围,这导致该时期植物群的总体发育较差。但是,在三叠纪晚期,喜欢水的植物,例如番茄脚纲植物(现在仅由俱乐部苔藓代表的维管植物),马尾和蕨类植物的出现,表明干旱的气候变为季风性更湿润的气候。皮带延伸至北纬60°N。亚热带至温带欧亚植物区系在约15°至60°N之间的一条带中,而在该带的北部是温带西伯利亚(安加拉)植物区系,延伸到三叠纪北极的10°以内。在南部大陆,适应凉爽,潮湿条件的二叠纪Glossopteris和Gangamopteris种子蕨植物区系被以Dicroidium为主的三叠纪植物区系取代,Dicroidium是首选温暖,干燥条件的种子蕨类植物,这表明二叠纪-三叠纪的主要气候变化边界。Dicroidium是蕨类植物的一个属,是南非南部晚三叠世莫尔特诺组和其他地区发现的冈瓦南古植物的一部分。该古植物从30°延伸到60°S以下。在15°N和30°S之间的赤道带,三叠纪几乎没有化石残留。
在海洋中,重要的一群仍活着的海洋浮游藻类在三叠纪晚期首次出现,而鞭毛藻在三叠纪晚期和侏罗纪早期经历了快速的多样化。整个三叠纪都存在着Dasycladacean海洋绿藻和蓝细菌。









美国第一夫人")


